1.2. Viljakust ja piimakust mõjutavad tegurid
Viljakusnäitajad - pesakonna suurus, pesakonnamass ja säilivus võõrutamisel - on suure majandusliku tähtsusega (Yen jt., 1987).
M. Varley ja R. Stedman (1994) on määranud emise näitajate osatähtsuse pesakonna suurusele (joonis 3), kust nähtub emise isiklike omaduste oluline mõju viljakusele. Eksperiment eelduseks oli, et suure varieeruvusega isiklike tunnuste mõned algandmed võivad selgitada olulist osa sigade viljakuse üleüldisest variatsioonist.
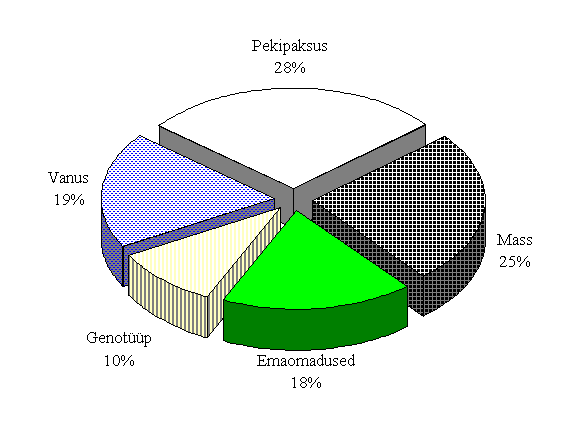
Joonis 3. Emise näitajate mõju pesakonna suurusele (Varley ja Stedman, 1994)
Üks tähtsamaid viljakust ja piimakust mõjutavaid tegureid on kindlasti põrsa sünnimass. L. Rydhmer (1992) leidis fenotüübilise ja geneetilise korrelatsiooni pesakonna suuruse ja põrsamassi vahel tiheda seose. Leiti, et pesakonna suurenemine on negatiivselt seotud põrsa-massiga. Suuremaid pesakond iseloomustab tendents, et sünnivad kergemad põrsad kui väiksemates pesakondades, niisiis võib aretus suurete pesakondade suunas viia suremuse suurenemiseni.
J.C. Kerr ja N.D. Cameron (1995) leidsid, et kui pesakonnas sünnib üks põrsas rohkem väheneb põrsaste võõrutuseelne suremus proportsionaalselt 0,12 põrsa võrra kuni põrsaste arv pesakonnas võrdub viiega, seejärel aga pesakonna suurenedes ühe põrsa võrra tõuseb ka põrsaste suremus proportsionaalselt 0,01 põrsa võrra (joonis 1).
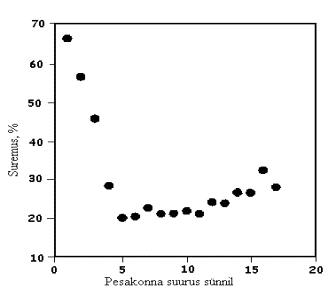
Joonis 1. Põrsaste võõrutuseelse suremuse sõltuvus pesakonna suurusest
(Kerr ja Cameron, 1995)
Väikestes pesakondades sündinud põrsad on suuremad, tugevamad ja elujõulisemad (Rydhmer, 1992; 1993). Nad haigestuvad vähem ja kasvavad kiiremini, mistõttu eriti suurte (nõrkade põrsastega) pesakondade tootmine ei ole majanduslikult õigustatud. Tootmisfarmides võiks eesmärgiks seada 10…11 põrsast pesakonnas, aasta keskmisena peaks emiselt saama vähemalt 20 põrsast (Meisner, 1990; Kersten, 1995).
J.C. Kerr ja N.D. Cameron (1995) on oma katsete põhjal koostanud graafiku põrsaste surevuse sõltuvuse kohta sünnimassist (joonis 2). Põrsaste suremus väheneb kiiresti põrsa sünnimassi suurenedes. Põrsamassi korral üle 1,5 kg on suremus on alla 10%.
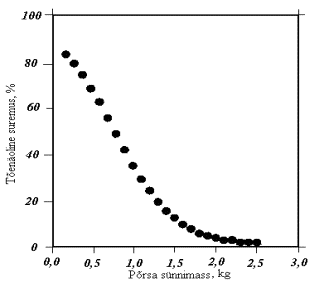
Joonis 2. Põrsaste võõrutuseelse suremuse sõltuvus sünnimassist (Kerr ja Cameron, 1995)
Oma töös on M.H. Fahamy ja C. Bernard (1971) põhjalikult käsitlenud pesakonna suuruse ja põrsaste elujõulisuse seost. Elusaltsündinud põrsaste suur arv ei taga veel, et suur arv põrsaid ka võõrutatakse. Põrsa sünnimass on aga elujõulisusega positiivselt seotud just esimeste nädalate kestel. Seega tuleb põrsaste arvu kõrval arvestada ka põrsaste sünnimassiga.
Ka H.E. Nielsen (1975) väidab, et suuremas pesakonnas väheneb tugevasti põrsaste elujõulisus põrsaste väiksema massi tõttu. Tabelist 5 on näha, et põrsaste suremus on kõige suurem siis, kui põrsa sünnimass on alla 1,20 kg.
Tabel 5. Põrsaste suremuse sõltuvus sünnimassist (Nielsen, 1975)
|
Põrsa sünnimass, kg |
Põrsaste arv |
Suremus, % |
|
Kuni 0,90 |
123 |
56,1 |
|
0,91...1,20 |
416 |
20,4 |
|
1,21...1,40 |
550 |
11,3 |
|
1,41...1,60 |
637 |
9,1 |
|
1,61...1,80 |
431 |
6,5 |
|
1,81...2,00 |
159 |
6,9 |
|
Üle 2,00 |
46 |
8,7 |
Vaatamata pesakonna suuruse negatiivsele seosele teiste viljakusnäitajatega on üheks aretuseesmärgiks suurte pesakondade saamine võõrutamisel. Elusaltsündinud põrsaste arv on üheks selektsiooni programmi valikutunnuseks Rootsis ja paljudes teistes maades (Brandt, 1990; Webb, 1991; Estany jt., 1993; Rydhmer, 1993; Sehested ja Ianssin, 1993). Sündinud põrsaste arvu asemel võõrutatud põrsaste arvu valiku aluseks võtmast takistab arvatavasti suhtumine, et sündinud põrsaste arv sõltub emisest, hiljem aga mõjutab ka farmer põrsaste säilivust. Et põrsad on esimeste elunädalate jooksul väga tundlikud keskkonnatingimuste muutuste suhtes, siis peab farmer tagama soodsad tingimused põrsaste säilimiseks ja arenguks. Põrsaste arvu kasvuga pesakonnas suureneb ka suremus, seega on geneetiline edu vähe väärt, kui see ei kajastu võõrutatud põrsaste arvu suurenemises. Seepärast on pesakonna suurus võõrutamisel parem valikutunnus kui pesakonna suurus sünnil, sest parem põrsaste säilivus näitab nende suuremat elujõulisust antud tingimustes. Siiski, kui kasutatakse ristandeid ja võõrutusvanus varieerub, on raske kehtestada standardit pesakonna suurusele võõrutamisel (Rydhmer, 1993).
Ristamise mõju hindamisel sigade viljakusele tuleb arvestada võrreldavate rühmade suurust. Katsetes on sigade arv sageli väike, mis ei võimalda teha objektiivseid järeldusi uurimistulemustest. Pig Improvement Company (1983) poolt on esitatud pesakondade miinimumarvud ja keskmiste 95% tõenäosusega erinevused (tabel 6).
Nendest nähtub, et objektiivsete järelduste tegemiseks peab pesakondade arv võrreldavates rühmades olema küllaltki suur. Siit võib ka selgust tuua küsimusele, miks mõned katsetulemused ei ühti teiste katsetulemustega.
Tabel 6. Pesakondade miinimumarvud ja keskmiste erinevus (Pig Improvenent Company, 1983)
|
Pesakondade arv kummaski rühmas |
Rühmade keskmiste erinevus, korda |
|
25 |
1,58 |
|
50 |
1,02 |
|
100 |
0,79 |
|
150 |
0,64 |
|
250 |
0,50 |
|
500 |
0,35 |
Väikeste rühmadega saab kergemalt hinnata siis, kui need on läbi viidud mitmes korduses ja tulemused on ühesuunalised. Edaspidi tuleks sigade viljakust käsitlevates uurimustes suuremat tähelepanu pöörata võrreldavate rühmade suurusele.
.2.1. Pärilikud tegurid
Seakasvatuse efektiivsust saab parandada kahel viisil:
1. parandades genotüüpi
2. parandades keskkonda ehk mittegeneetilisi tegureid.
Genotüüp seab lae, mil määral fenotüüpi saab parandada keskkonnategurite, pidamise ja söötmise parandamisega, aga alati on tähtis taotleda sigade geneetilise võime parandamist (English jt., 1988).
Paljud uurijad on püüdnud kindlaks teha geneetilise edu puudumise põhjusi pesakonna suuruses nii aretuskarjades kui ka valikueksperimentides (Skjervold, 1979; Johansson, 1981; Southwood ja Kennedy, 1991; Bolet jt., 1989). Mitmed kaasaegsed uurimistulemused, mis baseerusid viljakate emiste pesakonna andmetele, näitasid, et valik suuremate pesakondade järgi võib olla edukas (Legault, 1985; Bichard ja David, 1985). Tänapäeva teadlased on optimistlikud pesakonna suuruse geneetilise potentsiaali suhtes (Avalos ja Smith, 1987; Haley jt., 1988). A.J. Webb (1991) konstateerib, et on praktiline tegelikkus geneetiliselt suurendada pesakonda aastas 0,2…0,3 põrsa võrra.
Väike geneetiline edu viljakuses, eriti pesakonna suuruses, on saavutatud kaasajal (Kaplon jt., 1991; Southwood ja Kennedy, 1991). Võrreldes metssigadega on toimunud oluline muutus (Aumaitre jt., 1982). Väikese valiku edu põhjuseks pesakonna suuruses pole variatsiooni madal tase, sest tõugudevaheline geneetiline variatsioon on suur. Hiinast pärit meishani tõu kõrge viljakus on potentsiaali sobivaks näiteks, kuid pesakonna suuruse päritavus on sellest hoolimata madal (Bidanel, 1988). 1970. aastatel tõstatasid G.S. Strang ja J.W.B. King (1970) küsimuse pesakonna suuruse aretusväärtusest. Sellel ajal põhines peaaegu kogu valik toodangunäitajatel, peamiselt massi-iibel ja tailiha osatähtsusel. Nende tunnuste osas on saavutatud suurt edu (Lundeheim ja Eriksson, 1984) ja paljud uurijad eeldavad et tailiha optimum saavutatakse peatselt (Ollivier jt., 1990). Pealegi on selektsioon sega-mudelitega suurendanud lootust, et geneetilist edu võib saavutada tunnuste osas, millel on madal päritavus ja mida on mõjutavad rohkem keskkonna tingimused (Long jt., 1991). Pesakonna suurust on käsitletud näitajana, millel on suur majanduslik väärtus (Legault, 1978; Smith jt., 1983). See on samuti ainus viljakusnäitaja, mis on kantud paljudesse aretusprogrammidesse (Rydhmer, 1993). Tänapäeval on paljud aretusfirmad ja tõuraamatud seadnud prioriteediks suurendada viljakusnäitajate osa valikul.
Ebasoodne geneetiline korrelatsioon viljakuse ja teiste valikus kasutatavate näitajate vahel, keskkonna mõju ja nõrk geneetiline korrelatsioon esimese pesakonna viljakuse ning eluaja viljakuse vahel võivad olla viljakuse tähtsust vähendavad faktorid. Enamik teadlasi on seisukohal, et otstarbekam on parandada viljakusnäitajaid ristamiskombinatsioonidega, saavutades edu soovitud tunnustel ja täiendavalt manipuleerida keskkonnategurite ning majandamisega. Selline valiku ja pidamise koosmõju arvestamine kehtib ka lihakehanäitajate parandamisel (Johansson, 1981). Näiteks leidsid G. Bolet jt. (1984) negatiivse geneetilise seose lihasuse ja pesakonna suuruse vahel, mis on üks viljakust vähendavaid tegureid. L. Rydhmer (1993) seevastu leidis soodsa geneetilise korrelatsiooni lihasuse ja pesakonna suuruse vahel. Ka O. Vangen (1980) leidis, et valik suure toodangu suunas võib olla seotud pesakonna suuruse väikese kasvuga sünnil. Kontrastiks sellele leidsid K. Johansson ja B.W. Kennedy (1983) ebasoodsa geneetilise korrelatsiooni pekipaksuse ja pesakonna suuruse vahel. Vastupidiselt neile leidis L. Rydhmer (1993) ebasoodsa geneetilise korrelatsiooni kasvukiiruse ja pesakonna suuruse vahel. D.J.A. Cole (1971) andmetel pole aga ilmnenud tõendeid negatiivsete geneetiliste seoste kohta viljakuse ja teiste majanduslikult kasulike tunnuste vahel. P.W. Ferguson jt. (1985) leidsid positiivse geneetilise korrelatsiooni emise kehamassi ja pesakonna suuruse vahel. Soodsa korrelatsiooni massi-iibe ja 3-nädalase pesakonna suuruse vahel leidsid K. Johansson ja B.W. Kennedy (1983), ehkki see oli nõrgem võrreldes pesakonna suurusega sünnil. Põrsamass pole positiivselt seotud mitte ainult elujõulisusega, vaid kogu eluaja jõudlusega (Rydhmer, 1993). Geneetiline varieeruvus põrsa sünnimassis on suur. Järelikult näib kasulik olevat seada ka see aretuseesmärgiks.
L.D. Young jt. (1976) leidsid uurimuses soodsa geneetilise korrelatsiooni toodangunäitajate ja põrsamassi vahel. Emiste valikul kõrge toodanguindeksi järgi suureneb põrsa sünnimass (Vangen, 1980) ning saadakse suuremad sead (Jacques ja Vangen, 1987). R.M. McKay (1990) võttis valiku aluseks suure massi-iibe ja lihasuse seitsmes põlvkonnas, aga ei leidnud olulist seost emise kehamassi ja pesakonna suuruse vahel. On tõsiasi, et ühest viljakama tõu - meishani emised on üsna väikesed, mis näitab, et suur kasv ei ole suurte pesakondade saamiseks vajalik (Bidanel jt., 1990). Seoseid massi-iibe, keha mõõtmete ja pesakonna suuruse vahel on üldistavalt käsitlenud F.D. Brien (1986), märkides, et tulemused sigadel olid vastuolulised. K. Johansson (1981) ja L. Rydhmer (1993) on seisukohal, et vastuolusid erinevates uurimustes viljakuse ja toodangunäitajate vahel peab käsitlema ettevaatlikult, sest tunnuste definitsioonid ja keskkonnatingimused erinevad uurimustes suuresti.
Seakasvatusettevõtetes kasutatakse sageli suurema kasumi saamiseks heteroosi, sest ristandloomade jõudlus ületab nende puhtatõuliste kaaslaste keskmisi näitajaid. Tõugude ristamisel tekib heteroos, kui ristandite jõudlus on suurem puhtatõuliste vanemate keskmisest (joonis 4). Ristandite jõudlus võib olla suurem või väiksem kui parima puhtatõulise vanema oma, sõltudes ristatavatest tõugudest (Clutter jt., 1990).
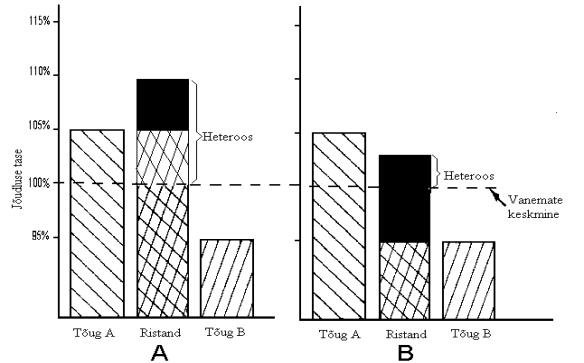
Joonis 4. Heteroosi erinev käsitlus. Ristandjärglaste jõudluse üleolek ühe (B) või mõlema (A) vanema jõudlusest (Clutter jt., 1990)
Heteroos on suurem madala päritavusega tunnustel, nagu pesakonna suurus, pesakonna võõrutusmass ja põrsaste säilivuse tase (tabel 7).
Tabel 7. Heteroosi tase ristamisel üle vanema keskmise (Webb, 1994)
|
Näitaja |
F1 % |
F2 % |
|
Pesakonna suurus sünnil |
8 |
3 |
|
Pesakonna suurus võõrutamisel |
11 |
6 |
|
Pesakonna võõrutusmass |
10 |
12 |
|
Võõrutusjärgne massi-iive |
0 |
6 |
|
Söödaväärinduse tase |
0 |
3 |
|
Lihakeha koostis |
0 |
0 |
Kui emasloom ja tema järglased on ristandid, võib pesakonna suuruse heteroos võõrutamisel kasvada 1,5 põrsa võrra (17%) ja pesakonna võõrutusmass 22% (Webb, 1994).
Ristamine ei muuda populatsioonis esinevaid geene, vaid sobitab neid palju soodsamatesse kombinatsioonidesse (Buchanan jt. 1990).
Ristamise heteroos ja sugulusaretuse depressioon ei avaldu ühesuguselt kõikidel jõudlusnäitajatel. Intensiivsemalt ilmneb see reproduktsiooninäitajatel ning peaaegu puudub lihajõudluse näitajatel. Seega on ristamise efekt madala päritavusega tunnustel suurem kui kõrge päritavusega tunnustel. Viimaste parandamiseks on aga paremad võimalused valiku abil (Meisner, 1985).
Tabelis 8 on antud jõudlusnäitajad, mida on võimalik mõjutada ristamise abil heteroosi kaudu.
Tabel 8. Heteroosi hinnangud sigade toodangunäitajatele (English jt., 1988)
|
Näitaja |
F1 jõudluse ülekaal vanemate keskmisest |
Täiendav heteroosiefekt ristandemiste kasutamisel |
||
|
|
diferents |
% |
diferents |
% |
|
Sündinud põrsaste arv, põrsas |
+0,30 |
3 |
+0,75 |
8 |
|
Põrsaste arv võõrutamisel, põrsas |
+0,45 |
6 |
+0,85 |
11 |
|
Põrsa võõrutusmass, kg |
+0,5 |
5 |
0 |
0 |
|
Pesakonna võõrutusmass, kg |
+9 |
12 |
+8 |
10 |
|
Võõrutusjärgne massi-iive, kg/päevas |
+0,04 |
6 |
0 |
0 |
|
Söödaväärinduse tase, kg sööta/kg kehamass |
-0,08 |
3 |
0 |
0 |
Tabelist on näha viljakuse, elujõulisuse ja piimakusega seostuvad tunnused, kusjuures heteroosiefekt on väiksem söödakasutuses, lihakehanäitajatel puudub aga hoopis. Need tunnused on kõrge päritavusega ja seetõttu on neid võimalik parandada valikuga. See annab aretajale võimaluse parandada valikuga paremini päritavaid tunnuseid ja rakendada heteroosi parandamaks madala päritavusega, kuid olulisi tunnuseid (English jt., 1988).
Arvestades sellega, et ristamise positiivne mõju on suurem madala päritavusega tunnustele, siis peaks ristamine olema efektiivne eeskätt reproduktsioonijõudluse parandamiseks (Meisner, 1985).
Valikul viljakusnäitajate järgi tuleb arvestada peale päritavuse ka genotüübi ja keskkonna vahelisi seoseid ning toodangunäitajate majanduslikku väärtust (Rydhmer, 1993).
1.2.1.1. Emise ja pesakonna tõulisus
Sigade aretuse ajalugu näitab, et teadlikud aretajad on kasutanud mõlemat - nii ristamist kui ka valikut, sest ristamisega suurendatakse populatsiooni heterogeensust, valikuga aga homogeensust. Heterogeensuse suurendamine annab võimalusi valiku efekti tõstmiseks. Seega tingivad nimetatud aretusvõtted teineteise vahelduvat kasutamist. Näiteks Taani seakasvatajad saavutasid edu kohalike sigade ja inglise suurt valget tõugu sigade ristamisel ning hiljem läksid üle puhasaretusele eesmärgiga veelgi parandada lihajõudlusnäitajaid. Niisugune ühesuunaline valik nõrgendas aga sigade konstitutsiooni ja vähendas elujõulisust ning kogu jõudlust. Seepärast hakati kasutama sigade ristamist. Enamik kultuurtõuge on saadud ristamise teel, millele on järgnenud puhasaretus (Rydhmer, 1993).
On korraldatud hulgaliselt sigade ristamiskatseid, milles on kasutatud kõikvõimalikke tõuge ja erinevaid ristamise variante. Tarberistamise kasutamine seakasvatuses on osutunud enamasti efektiivseks. Üksikutel juhtudel ei ole ristamine sigade jõudlust suurendanud (Meisner, 1985).
Tõul on suur mõju sigade viljakusele. Ristamisel on sigade sigivuse tulemused enamasti sõltunud sellest, milline on ristamiseks kasutatud tõugude viljakus. Seatõugude viljakusest mõnedes Euroopa riikides annab ülevaate tabel 9.
Tabel 9. Seatõugude viljakus eri riikides (Meisner, 1990)
|
Riik |
Tõug |
Põrsaste arv |
Riik |
Tõug |
Põrsaste arv |
|
Suurbritannia |
suur valge |
10,8 |
Holland |
hollandi landrass |
11,8 |
|
|
keskmine valge |
9,7 |
|
jorkšir |
11,7 |
|
|
berkšir |
8,7 |
|
pieträän |
9,7 |
|
|
tämvors |
8,0 |
Austria |
parandatud valge |
12,7 |
|
Saksamaa |
inglise |
11,4 |
|
landrass |
11,0 |
|
|
pikakõrvaline |
11,3 |
Šveits |
rootsi landrass |
10,8 |
|
|
lühikõrvaline |
11,0 |
|
suur valge |
10,8 |
|
|
pieträän |
10,5 |
USA |
jorkšir |
10,8 |
|
Prantsusmaa |
suur valge |
10,5 |
|
djurok |
9,8 |
|
|
prantsuse landrass |
10,2 |
|
taani landrass |
9,7 |
|
|
pieträän |
9,6 |
|
hämpšir |
8,7 |
|
Belgia |
suur valge |
11,0 |
|
poola-hiina |
8,0 |
|
|
belgia landrass |
10,0 |
|
berkšir |
7,7 |
|
|
pieträän |
9,6 |
|
tämvors |
7,4 |
Nagu nähtub on sigade viljakus riikides erinev. See võib olla tingitud erinevatest pidamistingimustest ja viljakuse hindamise metoodikast. Üheks viljakamaks tõuks on suur valge seatõug või tema sugulased. Suure viljakusega on veel landrassi seatõug, väikese viljakusega on tämvorsi, berkširi ja pietreeni tõug Euroopas ning Ameerika Ühendriikides. Üldiselt on Ameerika Ühendriikides aretatavad seatõud väiksema viljakusega kui Euroopa omad ning ka seal on väiksema viljakusega tämvorsi ja berkširi tõud, kohalikest tõugudest aga poola-hiina seatõug. Teistest märgatavalt suurema viljakusega on jorkširi seatõug.
Eeltoodust selgub tõugude erinev viljakus, kusjuures viljakamateks võib pidada suurt valget tõugu sigu, keda mõnedes riikides nimetatakse ka jorkširiks (Meisner, 1990).
Ka E. Meisneri (1985) poolt tsiteeritud paljud uurijad on katsete põhjal hinnanud üheks väärtuslikumaks tõuks suurt valget tõugu, kes ristamisel parandab nii reproduktsiooni- kui ka nuuma- ja lihajõudlust. Lisaks on täheldatud, et suurt valget tõugu sead on hästi vastupidavad tööstuslikes tootmistingimustes ning on vähemtundlikud stressifaktorite suhtes. Viljakad on ka landrassi ja djuroki tõugu sead. Nende omavaheline ristamine suurendab järglaste reproduktsioonijõudlust veelgi (Meisner, 1990). Võrreldes puhtatõulistega valmivad ristandid kiiremini, kasutavad kehamassi juurdekasvuks vähem sööta ja toodavad suurema kalorsusega rasvasemat liha (Bradford jt., 1953; Reinhold, 1938).
H.F. Yen jt. (1987) uurisid mitmete tegurite mõju jorkširi, djuroki, valge tsesteri ja hämpširi tõugu emiste reproduktsioonijõudlusele ning järeldasid, et landrassi ja jorkširi emiste pesakonnad on sünnil teistest suuremad ja 21 päeva vanuselt ka raskemad.
Üks jõudlustulemusi mõjutavaid tegureid on ristamiseks valitud tõugude jõudlusvõime ja selle pärandumine. Efektiivsem on parematesse liinidesse ja tõugudesse kuuluvate loomade ristamine. Seega tuleb ristamiseks valida nii tõuge kui ka üksikuid loomi (Meisner, 1990). Ristamine on geneetiliselt erinevate tõugude üksikisendite seemendamine. Ristamise eesmärgiks on kujundada seatüüp, kes on paremini kohanenud erinevate keskkonnatingimustega ning on paremate geneetiliste eeldustega kui mõni puhtatõuline (Rae, 1952).
Ristamise tulemused sõltuvad samuti suurel määral sellest, millist ristamismeetodit kasutatakse. Aretusmeetoditest on kasutatud lihtsat tarberistamist kahe tõu ning vahelduvat ristamist kahe või mitme tõuga. Kahe tõu lihtne tarberistamine on olnud efektiivne järglaste elujõu ja nuumajõudluse parandamisel. Mitmete autorite andmeil ei ole selle aretusmeetodi kasutamisel siiski saadud olulist efekti. Viljakuse suurendamise eesmärgil on viimasel ajal pööratud suuremat tähelepanu ristandemiste kasutamisele (Meisner, 1964; Cosic, 1978).
Kahe tõu mõju reproduktsioonijõudlusele on uurinud M. Milojic jt., (1978) ning H. Cocic (1978). Nad leiavad, et kahe tõu ristamine ei mõjuta oluliselt sigade reproduktsioonijõudlust. A.H. Kaщeнкo (1958) on täheldanud, et suurt valget tõugu emiste ristamine teiste tõugude kultidega vähendas emiste sigivust.
E. Meisneri (1964) katsetest aastail 1961…1963 Sootaga sovhoosis selgus, et suurt valget tõugu emiste paaritamisel eesti peekoni tõugu kultidega ei saadud poegimisel suuremaid pesakondi, kuid saadi raskemaid ja elujõulisemaid põrsaid, mille tulemusena ristandpesakondade suurus võõrutamisel ületas puhtatõuliste pesakondade suuruse 0,8…0,9 põrsa võrra. Ristandpõrsaste sünnijärgne kehamass ületas puhtatõuliste põrsaste oma keskmiselt 120 grammi ehk 10,6% võrra. Ristandpõrsad kasvasid ka pärast sündi kiiremini kui puhtatõulised põrsad ja olid võõrutamisel raskemad. Heteroos, mis avaldus esimese põlvkonna ristandjärglaste suurenenud elujõus ja kasvukiiruses, jäi püsima ka teise põlvkonna ristanditel.
O. M. Hale ja K. Bondari (1986) uurisid pikema aja vältel sigade vahelduvat (rotatsioon) ristamist. Kasutati djuroki, hämpširi, jorkširi ja ka landrassi tõugu sigu. Sigade viljakuses heteroosiefekti ei täheldatud. 1966. aastast 1979. aastani ei suurenenud ka keskmine viljakus, mis oli 10 põrsast pesakonnas. Pesakonna suuruse ja põrsamassi vahel täheldati negatiivset seost. Pesakonna suurenemisega 1 põrsa võrra vähenes põrsa kehamass sünnil 24 grammi 1966. ja 1967. aastal ning 32 grammi 1971. ja 1974. aastal, võõrutamisel oli see aga väiksem vastavalt 120 ja 150 grammi. Veel leidsid autorid, et emise kehamassi suurenemisel 1 kg võrra suurenes põrsa sünnimass 2 grammi võrra. Pesakonna sünnimassi suurenemisel 1 kg võrra suurenes pesakonnamass võõrutamisel 2…3 kg võrra.
Põhjalikud uurimused viis läbi E. Meisner (1967) Tartu rajooni Sootaga sovhoosis 1961…1965. aastatel. Eesmärgiks oli selgitada vahelduva ristamise mõju suurt valget ja eesti peekoni tõugu sigade reproduktsiooni- ning järglaste nuuma- ja lihajõudlusele.
Tabel 10. Ristamise mõju sigade reproduktsioonijõudlusele Sootaga sovhoosis 1961…1965. a. (Meisneri, 1967)
|
Vanemate tõug |
Pesakonna |
Viljakus |
Pesakonna |
Kahe kuu vanuselt |
||||
|
emis |
kult |
nr. |
arv |
|
mass 1 kuu vanuselt kg |
põrsaste arv pesa-konnas |
põrsa keskmine mass kg |
pesa-konna mass kg |
|
ESV |
ESV |
1. |
30 |
9,97 |
58,95 |
8,27 |
15,01 |
124 |
|
|
|
2...3. |
22 |
10,91 |
56,16 |
8,64 |
12,86 |
111 |
|
ESV |
EP |
1. |
19 |
10,32 |
67,29 |
9,00 |
15,68 |
141 |
|
|
|
2...3. |
19 |
11,05 |
66,79 |
9,68 |
15,66 |
152 |
|
ESVxEP |
ESV |
1. |
17 |
11,06 |
76,82 |
10,12 |
16,15 |
163 |
|
|
|
2...3. |
12 |
12,50 |
65,19 |
9,67 |
17,64 |
171 |
|
ESVxEPxESVxESV |
EP |
1. |
8 |
10,75 |
72,69 |
9,38 |
16,41 |
154 |
Katsetulemustest selgus, et lihtsa tarberistamise korral suurenes emiste viljakus ainult 1,5…3,3% (tabel 10). Kui aga järglaste saamisel kasutati ristandemiseid (vahelduv ristamine), suurenes põrsaste arv pesakonnas 8…15% võrra. Ristandpõrsad olid ka elujõulisemad, mis ilmnes väiksemas suremuses ja intensiivsemas kasvus. Ristandpesakondade põrsaste arv oli võõrutamisel 0,7…1,8 põrsa võrra suurem kui puhasaretuse teel saadud pesakondades. Intensiivsema kasvu tagajärjel oli ristandpõrsaste keskmine mass võõrutamisel suurem kui puhtatõulistel põrsastel. Selgus veel, et ristamine ei paranda oluliselt järglaste nuuma- ega lihajõudlust.
Suurt valget tõugu emiste ristamisel eesti peekoni tõugu kultidega suurenes pesakonnamass ühe kuu vanuselt esimesel poegimisel 8,34 kg ja teisel poegimisel 10,63 kg võrra. Ka teisel kuul kasvasid ristandpõrsad puhtatõulistest kiiremini ning teise põlvkonna ristandpõrsaste kehamass oli esimese põlvkonna ristandpõrsaste kehamassist suurem. Ristandpõrsad olid kahe kuu vanuselt puhtatõulistest märksa raskemad.
Sigade ristamine kui aretusmeetod võeti Eestis laiemalt kasutusele 70ndatel aastatel vastloodud Gagarini-nim. NST Eksperimentaalses Sealihatootmise Kombinaadis (EKSEKO). Reproduktsioonijõudluse väljaselgitamiseks korraldati katse kõikide farmi emistega (Meisner, 1990). Tulemus näitas, et ristandemised olid viljakamad. Esimest korda poeginud emiste pesakondades oli keskmiselt 0,5 ning kaks ja enam korda poeginud emiste pesakondades 0,7 põrsast rohkem kui puhasaretuse teel saadud pesakondades. Kahe tõu lihtsal tarberistamisel suurenesid pesakonnad puhasaretusega võrreldes vähe - 0,2 põrsa võrra pesakonnas. Piimakus ja pesakonnamass kahekuuselt suurenes peamiselt põrsaste suurema arvu tõttu pesakonnas. Kahekuuste ristand- ja puhtatõuliste põrsaste kehamass oli praktiliselt võrdne. Nuumajõudluse väljaselgitamiseks kaaluti ja mõõdeti EKSEKO sigade lihakehasid pärast tapmist Võhma lihakombinaadis. Selgus, et tapajärgselt oli ristandsigade lihakehamass 30 kg võrra suurem kui puhtatõulistel sigadel. Seega võib arvestada, et nende ööpäevane massi-iive pärast võõrutusperioodi oli ca 50 grammi puhtatõuliste omast suurem. Pekipaksuses olulisi erinevusi ei täheldatud.
Üksikasjalisemalt on sigade ristamist uurinud A. Timmi (1988). Peale Eestis aretatavate tõugude on ta ristamiseks kasutanud ka sissetoodud djuroki tõugu. Nendestki uuringutest ilmneb ristandsigade heteroosiefekt. Suurem on see reproduktsioonijõudluses, ulatudes üksikutes katsetes 30% piiridesse. Ööpäevane massi-iive oli ristanditel suurem 5,8…14% võrra ja lihajõudluse näitajad sõltusid suurel määral kasutatavate tõugude lihajõudlusest.
A. Timmi uuris Kehtna Põllumajandusloomade Kunstliku Seemendusjaama imporditud djuroki tõugu kultide sobivust eesti tõugudega ristamiseks. Eesti peekoni nooremiste seemendamisel sama tõugu kultide spermaga saadi pesakonnas 6,88 põrsast, suurt valget tõugu kultide puhul 8,17 ja djuroki kultidega 8,25 põrsast pesakonnas. Samal ajal oli farmi keskmine 7,85 põrsast pesakonnas. Suurt valget ja eesti peekoni tõugu ristandemiste seemendamisel djuroki tõugu kultide spermaga saadi 9,82 ja eesti peekoni tõugu kuldi puhul 10,82 põrsast pesakonnas. Uuriti ka suurt valget ja eesti peekoni ning djuroki ja eesti peekoni tõugu ristandpõrsaste kasvu. Selgus, et djuroki ja eesti peekoni ristandpõrsad kasvasid imetamisperioodil aeglasemalt, kuid pärast võõrutamist kiiremini kui suurt valget ja eesti peekoni tõugu ristandpõrsad. Võõrutamisel oli esimeste kehamass 6,4 ja teistel 6,6 kg, realiseerimisel aga vastavalt 27,8 ja 25,8 kg. Suurt valget ja eesti peekoni tõugu sigade ristandpesakondades oli keskmiselt 10,7 ning djuroki ja eesti peekoni tõugu sigade pesakondades aga 11,2 põrsast.
USAs ja Kanadas läbiviidud ristamiskatsed näitasid samuti heteroosiefekti - pesakond suurenes keskmiselt 0,87 põrsa võrra (Quintana, 1979).
G. Rali jt. (1978) poolt Rootsis tehtud ristamiskatsetes (landrass x jorkšir) saadi pesakonna suuruse efektiks 0,33 põrsast, tagasiristamisel 0,55 ja veelkordsel tagasiristamisel 0,42 ning mittesüstemaatilisel ristamisel 0,29 põrsa võrra parem tulemus kui puhtatõuliste vanemate puhul (tabel 11).
Tabel 11. Ristandemiste pesakondade suuruse üleolek puhtatõuliste
emiste pesakondadest (Ral jt., 1978)
|
Põrsaste arv |
|||
|
|
sünnil |
sündis elusalt |
3-nädalaselt |
|
Tarberistamine |
0,26*** |
0,27*** |
0,33*** |
|
Tagasiristamine |
0,55*** |
0,47*** |
0,55*** |
|
Veelkordne tagasiristamine |
0,40*** |
0,34*** |
0,42*** |
|
Mittesüstemaatiline ristamine |
0,30*** |
0,20*** |
0,29*** |
J. Wiebel'i (1977) andmetel suurendab ristamine viljakust 5,1%, põrsaste arvu 21 päeva vanuselt 6,0% ning põrsamassi 56 päeva vanuselt 6,3% võrra. G.W. Sherritt ja D.E. Younkin (1973) kirjutavad, et noortel ristandemistel on viljakus 8,5 ja puhtatõulistel 7,5 põrsast. Ristandpõrsad olid 8-nädalaselt 2 kg võrra puhtatõulistest raskemad.
D.L. Kuhlers ja S.B. Jungst (1987) kasutasid landrassi emiste ristamiseks djuroki, hämpširi ja jorkširi tõugu kulte. Esimese põlvkonna ristandemiste seemendamisel kasutati lähtetõu kulti. Selgus, et tagasiristamisel saadud emiste jõudlus oli sünnil 0,3 ja võõrutamisel 0,4 põrsa võrra väiksem kui esimese põlvkonna ristandemistel ning pesakonnamass 21 ja 56 päeva vanuselt 2,3 ja 10,9 kg võrra väiksem.
Oklahoma Riiklikus Ülikoolis uuriti kahe ja kolme tõu ristamise mõju sigade jõudlusele (Johnson jt., 1978). Kasutati djuroki, hämpširi ja jorkširi tõugu sigu. Ristandemistel sündis pesakonnas 0,93 ("0,32) põrsast rohkem ja võõrutamise ajal oli neid 1,24 ("2,7) võrra rohkem kui puhtatõulistel.
Hämpširi, jorkširi ja djuroki tõuga korraldas ristamiskatsed B. Ahlschwede (1979). Autor leidis, et ristamine parandab emiste jõudlust. Ristamisel sündis ja võõrutati rohkem põrsaid kui puhtatõulistel. Ristandsead olid elujõulisemad ja neid oli võimalik realiseerida nooremalt.
Prantsuse autorid (Legault jt., 1975) väidavad, et ristandemistel on pesakonnad sündimisel 8% ja võõrutamisel 12…15% võrra suuremad kui puhtatõulistel emistel. Samad autorid märgivad veel kuldi olulist mõju pesakonna suurusele. N.D. Dzapo (1978) peab oluliseks tõu mõju ristamisel. Tema uurimustes olid aga belgia ja saksa landrasside ristandid vähem vastupidavad kui puhtatõulised eakaaslased. P. Jensen (1978) on avaldanud ulatusliku uurimuse sigade ristamise kohta, kus kasutati taani landrassi, jorkširi, norra landrassi, hämpširi ja rootsi landrassi. Sigade viljakus sõltus suurel määral kasutatud tõugudest. Suremuse erinevus oli kuni 1,7 põrsast pesakonna kohta.
Rootsis korraldasid H. Simensson ja K. Andersson (1978) ristamiskatsed, kus landrassi ja jorkširi ristandemised seemendati puhtatõuliste landrassi, jorkširi või hämpširi kultidega. Hämpširi tõugu kuldid vähendasid põrsaste arvu pesakonnas. Kui landrassi ja jorkširi ristandemiste seemendamisel kasutati landrassi tõugu kulte, saadi 10,19 põrsast pesakonnas, hämpširi tõugu kultide puhul aga 9,7 põrsast pesakonnas; jorkširi kultide kasutamisel oli pesakonnas 10,25 põrsast.
Ulatusliku uurimismaterjali sigade ristamistulemuste kohta on esitanud Kanada autorid W. B. Holtmann (1975), M. H. Fahmy (1975), M. H. Fahmy ja W. B. Holtmann (1977), M. H. Fahmy jt. (1978) ning B.W. Kennedy ja J. E. Moxley (1978). Võrreldi 8 tõu 28 erinevat ristamiskombinatsiooni. Uuriti nii reproduktsiooni-, kasvu- kui ka lihakehanäitajaid. Kõikide näitajate alusel osutus parimaks ristamiskombinatsiooniks hämpšir x landrass. Kui välja arvata reproduktsioonijõudlus, osutusid paremaks landrassi ja jorkširi ristandid. Kolme tõu ristamisel saadud järglased olid üldiselt paremad puhtatõulistest või kahe tõu ristanditest. Peaaegu võrdseid tulemusi andsid ka nelja tõu erinevad ristamisskeemid, kus kasutati nii ristandemiseid kui ka ristandkulte. M.H. Fahmy (1975) leidis, et erinevatel ristamiskombinatsioonidel oli tõenäoline mõju sigade kõigile reproduktsiooninäitajatele, välja arvatud tiinuse kestus. Ka V. Moskal (1978) pidas ristamist oluliseks aretusmeetodiks sigade reproduktsioonijõudluse parandamisel Tšehhoslovakkias, kus kõige paremaid tulemusi saadi kolme tõu ristamisel.
O.W. Robison (1972) on üldistanud hulgaliselt materjali emiste mõju kohta sigade aretuses. See annab tunnistust ema mõju jätkumisest pärast põrsaste võõrutamist, seda eriti kehamassile 140 päeva vanuselt, pekipaksusele, ovulatsiooni intensiivsusele ja pesakonna suurusele.
Inglise firma Pig Improvement Company (1983) poolt avaldatud andmetel on ristandemiste viljakus 0,75 põrsa võrra suurem kui puhtatõulistel suurt valget tõugu emistel. Nimetatud firma pöörab suurt tähelepanu sigade viljakusele ning püüab seda aretusvõtetega suurendada.
A.C. Clutter jt. (1990) leidsid, et puhtatõulise emise seemendamisel teise tõu kuldiga suureneb sündinud põrsaste arv pesakonnas oluliselt. Samas olid ristandpõrsad elujõulisemad kui puhtatõulised. Seega olid kahe tõu ristandpesakonnad võõrutamisel 11,8% raskemad kui puhtatõulised. Ristamise eelise maksimum realiseeritakse aga ristandemiste kasutamisel, kus võõrutatud põrsaste arv pesakonnas oli 8,7% suurem. Selline paranemine saavutati tänu elusaltsündinud põrsaste arvu kasvule (4,7%) ja põrsaste suuremale säilivusele. Ristandemiste pesakonnamass 21-päevaselt oli 29% suurem puhtatõuliste emiste puhtatõulisest pesakonnast.
Tõusiseselt on päritavus pesakonna suurusele, põrsa sünnimassile ja võõrutusmassile väga madal. G.S. Strang ja J.W.B. King (1970) on leidnud päritavuse hinnanguks suurel valgel tõul 3…11%. Järelikult nendel tunnustel võib loota ristamisel heteroosi, mida on väidetud ka H.T. Fredeeni (1957b) ja S. Skårman (1965) töödes. Kõige detailsem eksperiment viidi läbi Rootsis, kus S. Skårman (1965) võrdles suure valge ja landrassi sigade retsiprookse ristamise tulemusi puhtatõuliste sigade jõudlusega. Tema katsetes olid ristanditel suuremad pesakonnad sünnil, suurem elujõulisuse tase võõrutamisel ning suurem põrsa ja pesakonnamass kolme ja kaheksa nädala vanuselt (tabel 12).
Tabel 12. Ristandpesakondade jõudluse üleolek puhtatõulistest pesakondadest (Bitchard jt., 1971, järgi).
|
|
Ristandpesakond |
Ristandemis |
||||||
|
Tunnused |
(a) |
(b) |
(c) |
(d) |
(a) |
(b) |
(c) |
(d) |
|
Põrsaid pesakonnas, % |
|
|
|
|
|
|
|
|
|
sünnil |
2 |
7 |
5 |
-11 |
5 |
9 |
10 |
7 |
|
võõrutamisel |
5 |
9 |
12* |
3 |
8 |
7 |
12* |
14 |
|
Pesakonnamass |
|
|
|
|
|
|
|
|
|
võõrutamisel, kg |
10 |
10 |
11 |
6 |
11 |
8 |
- |
17 |
* - võõrutati 42-päevaselt
(a) G.S. Smith ja J.W.B. King, 1964;
(b) S. Skårman, 1965;
(c) M.P.M. Vos, 1970;
(d) P.J. Cunningham jt., 1967.
Samalaadne töö Hollandis (Vos, 1970) kinnitas neid tulemusi. Suureulatuslik eksperiment Oklahomas (Cunningham jt., 1967) ei näidanud aga pesakonna suurenemist ristamisel, kuid kinnitas märgatavat mõju elujõulisusele ja pesakonna võõrutusmassile. Suurbritannias C. Smithi ja J.W. Kingi (1964) poolt läbiviidud katsed kinnitasid veelkord ristamise suuremat tulemuslikkust.
Kõik need andmed viitavad eelistele, mida annavad erinevate puhtatõuliste vanemate ristandjärglased. Edu võib oodata, kasutades ristandemiseid mõnede pesakonnanäitajate parandamiseks. Sellist edu pesakonna suurusele ja massile on saadud kõigis katsetes (v.a. Skårman, 1965).
Kokkuvõttes võib väita, et tõugudevaheline ristamine on üks võtteid, mis võimaldab suurendada sigade viljakust ja piimakust. Ristamiseks valitavad sead peavad olema väärtuslikud ja sigade nõuded söötmise ja pidamise osas rahuldatud. Viletsate sigade paaritamine ja ristandsigade pidamine kehvades tingimustes ei anna soovitud tulemusi (Meisner, 1990).
1.2.1.2. Päritavus
Enamik sigade majanduslikult kasulikke tunnuseid on mõjutatud ka pärilikkusest. J.G. Mendeli pärilikkuseseadused käsitlevad dominantseid ja retsessiivseid geene, kus kahe heterosügootse indiviidi kombineerimisel on fenotüübiline lahknemine 1. põlvkonnas (F1) 3:1 ja genotüübiline 1:2:1. Seda ei saa siiski aluseks võtta valikul majanduslikult kasulike tunnuste järgi. Enamik nendest tunnustest on määratud paljude geenide koostoimest, mistõttu karja parandamisel tuleb uurida nende mitmekordselt toimivate geenide koguefekti erinevate valiku- ja ristamisskeemide korral. Teoreetiliselt võib päritavus olla 0…1,0 piirides (Pond ja Maner, 1984).
Põhjalikud emiste viljakuse uurimused on andnud pesakonna suurusele madala päritavuse - h2 = 0,11 (Hill ja Webb, 1982; Johansson, 1981; Haley jt., 1988; Lamberson jt., 1991). Pesakonnamassi päritavus on veidi kõrgem (tabel 13). Pesakonna suurusel on leitud negatiivne kovariatsioon otsese geneetilise ja emaefekti vahel, eriti noorte emiste pesakondadel (Johansson, 1981).
Tabel 13. Ülevaade pesakonna suuruse päritavusest
|
|
Põrsaste arv |
Pesakonnamass |
||||
|
Autor |
elusalt sündinud |
3-nädalaselt |
8-nädalaselt |
3-nädalaselt (kg) |
8-nädalaselt (kg) |
|
|
Urban jt. (1966) |
0,08 |
- |
0,13 |
- |
0,19 |
|
|
Legault (1970) |
0,071 0,112 |
0,01 0,09 |
- - |
- - |
0,03 0,05 |
|
|
Strang ja King (1970) |
|
|
|
|
|
|
|
suur valge |
0,072 |
0,07 |
0,09 |
0,08 |
0,03 |
|
|
Strang ja Smith (1979) |
|
|
|
|
|
|
|
suur valge |
0,041 |
0,03 |
0,05 |
0,09 |
0,18 |
|
|
landrass |
0,092 |
0,10 |
0,06 |
0,11 |
0,03 |
|
|
|
0,071 |
-0,02 |
0,00 |
0,07 |
0,12 |
|
|
Johansson (1981) |
|
|
|
|
|
|
|
1. pesakond |
0,181 |
0,16 |
- |
0,21 |
- |
|
|
2. pesakond |
0,151 |
0,15 |
- |
- |
- |
|
|
Hill ja Webb (1982) |
0,12 |
- |
- |
- |
0,09 |
|
|
Haley jt. (1988) |
0,09 |
- |
- |
- |
- |
|
|
Lamberson jt. (1991) |
0,07 |
- |
- |
- |
0,06 |
|
|
Rydhmer(1993) |
0,13 |
- |
- |
- |
- |
|
1 - poolõvede analüüs
2 - tütar - ema analüüs
Põrsa keskmise sünnimassi päritavus on kõrge (h2=0,45) võrreldes teiste viljakusnäitajatega (Webb ja King, 1976; Rydhmer, 1993). Võib vaielda, kas keskmine põrsamass on emise viljakusnäitaja või mitte. Siiski leiti Rootsis nullilähedane põrsa sünnimassi päritavus (Rydhmer, 1992). C. Rodriques jt. (1993) leidsid 3-nädalase põrsamassile väga madala päritavuse. Et raskemad põrsad on elujõulisemad, peaks kasvukiirus olema positiivselt seotud võõrutatud põrsaste arvuga.
Kõrgema päritavusega tunnuseid on kergem parandada valikuga, kasutades geenide aditiivset toimet. Siiski, kui tunnus on madala päritavusega, aga samal ajal kõrge varieeruvusega, võib seda parandada just samavõrra kui tunnust, millel on kõrge päritavus, kuid väga madal variatsioonitase populatsioonis. Vastupidi, madala päritavusega tunnuseid mõjutavad keskkonnategurid tunduvalt rohkem, samuti toimivad mitmesugused geneetilised mitteaditiivsed mõjud, mida ei saa valikul rakendada. Sündinud põrsaste arvu, mille keskmine päritavus on 10%, saab valiku abil mõjutada 10 % ulatuses tema muutlikkusest. Pesakonna suuruse variatsiooni ülejäänud 90% on osaliselt mitteaditiivse toime, osalt aga keskkonnafaktorite mõju, millest ainult mõned on määratavad ja kontrollitavad. Mitteaditiivset mõju saab rakendada mõnedes hästi plaanitud ristamisskeemides, püüdes saavutada suuremat kontrolli keskkonnafaktorite üle, mis mõjutavad pesakonna suurust, nagu pidamistingimused, haigused ja söötmine. Mõnede seakasvatuse seisukohalt tähtsate tunnuste päritavus R. Englishi (1988) järgi on esitatud tabelis 14.
Tabel 14. Tähtsamate tunnuste päritavus sigadel (English 1988)
|
Tunnus |
Tase |
Piirid |
Keskmine |
|
Pesakonna suurus: sünnil |
madal |
0,05…0,15 |
0,10 |
|
võõrutamisel |
madal |
0,05…0,15 |
0,10 |
|
Põrsamass võõrutamisel, kg |
madal |
0,05…0,15 |
0,10 |
|
Põrsaste massi-iive, g/ päevas |
kõrge |
0,45…0,55 |
0,50 |
Kanada seaaretusspetsialisti Jim Morrise 18. 11. 1994. aasta loengu põhjal on R. Kaselo (1995) koostanud tabeli, kus on toodud erinevate tunnuste päritavus ja meetodid nende parandamiseks (tabel 15).
Nagu tabelist näha, mõjutavad pidamistingimused kõiki jõudlusnäitajaid. Liha kvaliteeti saame tõhusalt mõjutada aga vaid selektsiooni ja pidamistingimuste muutmisega. Pesakonna suuruse parandamisel on aga tähtsal kohal ristamine ja pidamistingimuste muutmine.
Tabel 15. Jõudlusnäitajate päritavus ja nende parandamise meetodid (Kaselo, 1995)
|
Näitaja |
Päritavus |
Parandamise meetodid |
||
|
|
|
selektsioon |
ristamine |
pidamistingimuste muutmine |
|
pesakonna suurus |
+ madal |
|
+ |
+ |
|
*massi-iive *söödaväärindus |
++ keskmine |
+ |
+ |
+ |
|
*rümba pikkus *seljapeki paksus *lihassilma suurus |
+++ kõrge |
+ |
|
+ |
T.J. Revelle ja O.W. Robison (1973) märkisid, et viljakusnäitajate päritavust on võimalik suurendada pesakonna suuruse standardiseerimisega.
1.2.2. Keskkonnategurid
Keskkonnatingimused võivad moonutada tõelist pilti looma geneetilistest eeldustest, sest genotüübi realiseerumist fenotüübina piiravad konkreetsed tingimused, milles organism areneb. Geneetiliselt kõrge jõudlusvõimega loom võib halbadesse pidamistingimustesse sattudes oma jõudluselt isegi alla jääda keskpärase geneetilise potentsiaaliga loomale. Katse ajal kehtivad keskkonnatingimused peaksid olema võimalikult lähedased nendele, mis kehtivad enamiku loomade pidamisel (Teinberg, 1978). Paljud spetsiaalselt koostatud genotüübi - keskkonna koosmõju katsed pole eriti ulatuslikud ja allikaks on sageli olnud juhuslikud vaatlused. Üks peamisi faktoreid, mis avaldab vastumõju ristamiseksperimentide teostamisel, on huvide konflikt, mis avaldub ulatusliku eksperimendi detailide planeerimisel. Näiteks soovivad söötmisspetsialistid enamasti uurida väga paljusid söötasid korraga ja eelistavad eksperimente, mis tekitab segaduse tõugude võimaliku koosmõju ja pesakonna efekti vahel. Aretajad teisest küljest soovivad sageli võrrelda erinevaid ristandeid ja on tavaliselt varustatud paljude arvuliste näitajatega, nii et on raske valida erinevate söödaväärtuste käsitluste vahel. Mõlema eriala teadlased peaksid laiendama materjali, millega nad töötavad ja arvestama keskkonnafaktoreid, et saada tõepärasemaid tulemusi (Cole, 1971).
1.2.2.1 Emise vanus
Uurimised näitavad, et rootsi suurt valget tõugu emiste pesakond suureneb kuni 5. poegimiseni (Tehver, 1953). Täiskasvanud emistel on suuremad pesakonnad ja põrsad sünnivad raskemana. Vanematel emistel on pesakonnad keskmiselt 15% suuremad kui esmapoeginud emistel (Schmidt ja Bretschneider, 1954).
H.R. Gaugler jt. (1984) ning B. Shostak ja B. Benkov (1992) leidsid emise vanuse olevat olulise (P<0,05), kusjuures esimesed pesakonnad olid väiksemad ja kergemad nii sünnil kui võõrutamisel.
Põrsaste suremuse uurimisel leidsid M.H. Fahmy jt. (1978), et esimeste pesakondade põrsaste suremus oli 19%, mis oli 2% võrra suurem kui teisel pesakonnal. Suremus sünnil oli teisel pesakonnal 2,3% kõrgem, kuid kolme elunädala jooksul madalam. Erinevused perioodide vahel ei osutunud tema andmeil oluliseks. N.C. Nielson jt. (1974) leidsid esimese ja teise pesakonna vahel 3,1%-lise erinevuse suremuses, kusjuures suremus oli suurem esimeses pesakonnas.
Põhjaliku ülevaate emiste vanuse mõjust nende tütarde viljakusnäitajatele on toonud H. Pärnamägi (1962). Võttes emise vanuse aluseks pesakonna numbri ja esimese pesakonna näitajad võrdseks 100%ga, leidis H. Pärnamägi protsendilised erinevused esimese pesakonna jõudlusest. Emiseperekondade vahel esines suuri erinevusi, mida tuleks valikul arvestada. Kõige suuremateks osutusid kolmas ja neljas pesakond. Viljakus püsis enam-vähem samal tasemel kuni üheksanda pesakonnani ning langes seejärel alla esimese pesakonna taseme. Seega olid kõige väiksemad esimene ja üheksas pesakond (joonis 5).
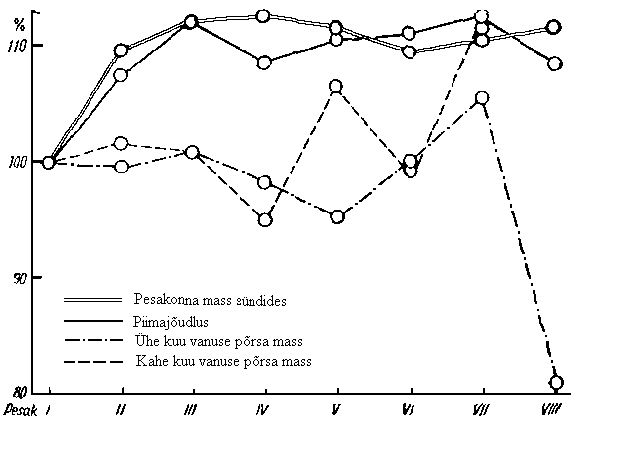 Joonis
5. Emise jõudluse olenevus nende ema vanusest (Pärnamägi,
1962)
Joonis
5. Emise jõudluse olenevus nende ema vanusest (Pärnamägi,
1962)
30 päeva piimajõudlus suurenes 3. pesakonnani, püsides edaspidi enam-vähem samal tasemel. Sarnaselt suurenes ka pesakonna sünnimass. Suurem oli piimajõudlus teise ja kolmanda pesakonna ajal ning madalaim esimese pesakonna ajal.
Ühe kuu vanuse põrsa keskmine mass oli kõige suurem seitsmendas ja kõige väiksem kaheksandas pesakonnas. Võrreldes esimese pesakonnaga järske tõuse järgnevatel pesakondadel (v.a. 7.) ei ole. Kahe kuu vanuse põrsa keskmine mass oli samuti kõige suurem seitsmendas pesakonnas.
K. Johansson (1981) leidis, et erinevate pesakondade suuruse päritavus on ligikaudu ühesugune - h2 = 0,15…0,17.
G. Bolet ja C. Felignes (1981) ja K. Johansson (1981) leidsid suhteliselt madala geneetilise korrelatsiooni erinevate pesakondade suuruse vahel. Sellel võib oodata peamiselt esimese ja teise pesakonna vahel. Nõrgal korrelatsioonil võib olla negatiivne mõju pesakonna suurusele, kui emiste valik toimub esimese pesakonna viljakusnäitajate järgi.
1.2.2.2. Emise söötmistase, Poegimissesoon
ja majand
Söötmise ja pidamisega ei saa muuta sigade pärilikkust ega tunnuste päritavust. Nende abil ei ole võimalik tõsta jõudlust üle geneetiliselt võimaliku piiri. Soodsate tingimuste loomisel on võimalik esile tuua või avastada looma geneetilist potentsiaali. Sellest ei tohi aga järeldada nagu poleks aretustöös vaja keskkonnatingimusi arvestada.
Mitmed E. Meisneri (1990) poolt tsiteeritud autorid väidavad, et ristamine on efektiivsem paremates söötmis-pidamistingimustes, mis on ka loogiline. Siiski on mitmed uurijad saanud ristandsigadega häid tulemusi ka viletsates söötmistingimustes. On täheldatud, et tarberistamine on efektiivne nii heades, rahuldavates kui ka halbades söötmis-pidamistingimustes. Samal ajal on mõned autorid väitnud, et ristamise efekt väheneb intensiivse nuuma tingimustes. Ristandsigadel on oma heterogeensuse tõttu eeliseid vähesobivates tingimustes. Samal ajal on aga viiteid selle kohta, et ristandsigade geneetilised eelised realiseeruvad rohkem paremates söötmis-pidamistingimustes (7@Db0>@&, 1977). Seega ei ole senini välja kujunenud üksmeelset seisukohta sigade söötmis-pidamistingimuste ja ristamisel ilmneva heteroosi koosmõju kohta sigade jõudlusele. Üldiselt on tulemused katsetingimustes siiski paremad kui tootmistingimustes. Nii võib tegelikkuses jääda mulje, et viletsates tingimustes jääb ristamisefekt nigelaks või puudub üldse (Meisner, 1985; 1990).
Sigade kasvule ja arengule avaldab suurt mõju aastaaeg ja majand. E. Meisner (1980) leidis, et kõigis majandites kasvasid talvel sündinud põrsad aeglasemalt kui suvel sündinud põrsad. Erinevus oli kõrge statistilise usutavusega (P<0,001) kõigil uuritud vanuseperioodidel. Samuti oli sigade kehamassis majandite viisi suuri tõenäolisi erinevusi. Sigade söötmis- ja pidamistingimused eri majandites ja tavaliselt ka ühes majandis eri aastaaegadel olid oluliselt erinevad. Ka H.R. Gaugler jt. (1984) leidsid, et sügisel sündinud pesakonnad on väiksemad ja massilt kergemad nii sünnil kui ka võõrutamisel. M.H. Fahmy jt. (1978) leidsid suremuse olevat kõrgema suvekuudel, mis aga erineb R.W. Pomeroy (1960) ja L. Bäckströmi (1973) poolt tsiteeritud paljude autorite töödest. See tuleneb ilmselt pidamistingimuste erinevustest eri maades. Külmadel kuudel on lauda sisetemperatuur konstantne ja säilitab palju ühtlasema taseme, aga suvekuudel on temperatuur palju kõikuvam, tõustes mõnel pool suhteliselt kõrgele. Kõrgete temperatuuride korral on imetavad emised põrsaste suhtes üldiselt vähem hoolivamad, mistõttu suureneb põrsaste suremus emise poolt tekitatud vigastustest (Fahmy jt., 1978).
Võrdsustades jaanuari viljakusnäitajad 100%-ga, leidis H. Pärnamägi (1962) teiste kuude erinevuse. Viljakus leiti olevat väikseim novembris ja detsembris poeginud emistel, suurenedes järsult veebruaris, märtsis ja aprillis. Seda peaks ära kasutama noortelt emistelt suuremate pesakondade saamiseks (joonis 6).
Veebruaris poeginud emiste 30 päeva piimajõudlus tõusis tunduvalt rohkem kui viljakus ja langes pisut koos pesakonna suuruse langusega märtsikuus. Aprillis ja mais poeginud emiste piimajõudlus langes tunduvalt rohkem. Suurim piimajõudluse tõus oli suve ja sügise algul, langedes novembris koos viljakusega kõige madalamale tasemele.
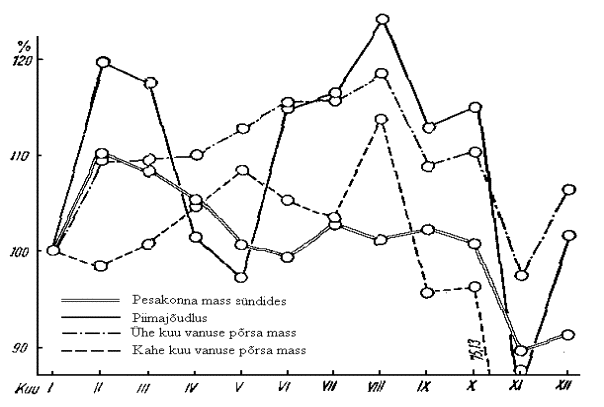
Joonis 6. Emise reproduktsioonijõudluse olenevus poegimiskuust (Pärnamägi, 1962)
Ühe kuu vanuse põrsa keskmine mass suurenes veebruaris vastavalt viljakusele ja pidevalt ka edasi kuni augustini olenemata viljakusest ja piimajõudlusest. Sealt alates langes nende mass järsult novembrini, et siis koos viljakuse ja piimajõudlusega uuesti tõusta.
Kahe kuu vanuse põrsa keskmine mass tõusis emise piimajõudlusest tunduvalt vähem ja langes novembris võrreldes jaanuariga. Siin olenes kahe kuu vanuse põrsamass ühe kuu vanuse põrsa keskmisest massist.
Kokkuvõtteks märgib H. Pärnamägi (1962), et kõige halvemad jõudlusnäitajad olid novembris ning kõige paremad veebruaris ja märtsis sündinud pesakondadel.
E. Meisner (1985) leidis, et kui katsetingimustes on sigade ristamine andnud paremaid tulemusi, siis on ristamis-efekt jäänud tagasihoidlikumaks. Nii on katsetingimustes saadud 10…20%-line ristamise efekt tootmistingimustes samasugust ristamisskeemi puhul tootmistingimustes kahanenud mõneprotsendiliseks.
Võrreldes sigu kasvu ja arengu järgi tuleb arvestada ka aastaaega ja majandi söötmis-pidamistingimusi.